

Shadows of Evolution
Shadows of Evolution

Charles Darwin’s theory, in its modern form, says that life has evolved over billions of years by natural selection working on genetic mutations. Evolutionists still debate the details of how this supposedly happened, and their suggestions have grown more sophisticated with the advance of science and technology. However, some arguments advanced in favor of the theory have not kept pace with these changes. They linger on in textbooks without being updated. Other ideas sound similar, but the evidence and reasoning have changed.
This article is a response to three ideas that persist either in updated or outdated forms. These are: vestigial organs; atavisms (or reversions); and embryonic recapitulation. All three are similar in suggesting that living things sometimes reveal remnants of their evolutionary past. Darwinists have used these arguments to make their case for the common origin of all species. As we will see, however, the same evidence often suggests common design by an intelligent Creator.
VESTIGIAL ORGANS
Charles Darwin cited several cases where animals have diminished or unused parts (1859, pp. 175-179). For example, the ostrich has wings but cannot fly, and cave fish have eyes but cannot see. Darwin used this as evidence that animals have changed over time: ostriches descended from birds with functional wings, and blind cave fish descended from fish with functional eyes. By the end of the nineteenth century, scientists had compiled a long list of such vestigial and rudimentary structures. In man, wisdom teeth, the coccyx, and the appendix were included as remnants of our animal past. Evolution, it was thought, eventually would eliminate these “useless” structures from our bodies.
However, with the advance of scientific knowledge over the last hundred years, it is difficult to find any “useless” body part. It now appears that vestigial simply meant “of unknown function.” Let us examine just three examples from the human body (summarized from Bergman and Howe, 1990).
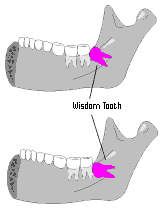 | 1. Some people experience problems with their wisdom teeth (or third molars). These teeth sometimes grow improperly, and have to be removed. Evolutionists explain this problem by saying that the human jaw is getting smaller. As apes we had the right number of teeth, they assert, but now there are too many for our modern jaw. However, many people have healthy, useful wisdom teeth. That they can cause problems may be a function of our changing diet. Or perhaps bad wisdom teeth represent a physical weakness, like failing eyesight, or hardening of the arteries. This does not suggest poor design, but a fallen state from perfect creation. |
 | 2. The coccyx is attached to the lower end of the vertebral column, and consists of three to five (usually four) fused vertebrae. Evolutionists believe this is a rudimentary tail, and it often is referred to as the “tail bone.” However, the coccyx serves a very real function as an anchor for muscles and tendons. These have a role in bowel movements, giving birth, leg movement, and other functions in the lower torso. |
 | 3. Perhaps the most popular example of vestigial structures is the appendix. This is a small, worm-shaped tube attached to the cecum at the end of the ascending colon. Many books on anatomy say that it has no function. Evolutionists have suggested that it was once part of a much larger cecum in our herbivorous ancestors. Like wisdom teeth, the appendix can cause problems. Sometimes it can become infected and, in severe cases, require removal. Like wisdom teeth, countless people have had an inflamed appendix removed with no negative, long-term effects. However, this does not mean that the appendix has no use at all. While all of its functions are not fully understood, the appendix has a rich supply of blood and contains masses of lymphatic tissues in its walls. This suggests that it may play a crucial role in the body’s immune system, especially in defense against infection in young children. Its presence may also boost antibody production in the spleen, and may play a role in preventing cancer. |
What about the diminished size of ostrich wings, and cave fish with eyes that cannot see? Darwin could not imagine why a Creator would design such imperfect creatures. However, Darwin made the assumption that these structures have no obvious purpose. Yet the ostrich wing is used in courtship displays, for balance during running, and to shield the nest from the Sun. Further, it is quite conceivable that degenerative changes could have occurred since the creation. For example, mutations may have caused the loss of sight in cave-dwelling animals, with no detriment to their survival. Even if these eyes are vestigial, they represent only limited change from the sighted condition (Frair and Davis, 1983, p. 29). Such vestiges do not support Darwin’s claim that lack of use is a driving force in large-scale evolution.
ATAVISM
The word “atavism” comes from the Latin atavus, which means father of a great-grandfather. French, and later British, doctors adopted it as a term to describe cases in which “a bodily peculiarity, deformity or disease, existing in a family, is lost in one generation; reappearing in that which follows” (Holland, 1839). In evolutionary theory, atavism (or reversion) refers to the appearance of features or traits belonging to a hypothetical ancestor.
Darwin gradually compiled a list of potential atavisms, but he settled on two main examples: the horse, and his favorite animal, the pigeon (1859, pp. 195ff.). In the case of the pigeon, he noted that a grayish-blue bird, resembling the ancestral rock pigeon, would appear occasionally among the progeny of exotic breeds. The case of the horse was not so simple because the ancestral form no longer exists. However, Darwin observed that zebra-like stripes often would appear on asses, quaggas, and many breeds of horses. “I venture confidently,” he wrote, “to look back thousands on thousands of generations, and I see an animal striped like a zebra” (1859, p. 201). For Darwin, this random appearance of a distinct trait among several varieties or species proved their common ancestry.
Today, the popular examples of atavism are more sensational and difficult to document. Perhaps the most commonly cited case is that of human babies born with a “tail.” This is supposed to represent a reversion to our animal ancestry. However, more detailed studies have shown that this caudal appendage, as it is known, contains no bone and is not connected to the vertebral column. It may be nothing more than excess tissue covering the lower end of the spinal column. This abnormality is no more a reversion than other birth defects such as spina bifida (see Gish, 1983).
So, how do reversions fit into evolution? We know from genetics that a characteristic can lay hidden for many generations. It may not appear in offspring unless both parents carry the gene for this characteristic. Or, a mixing of the parents’ genes at conception may recreate a former trait purely by chance. Either of these processes may cause “reversions,” but this is not the real issue.
Darwin wanted to show that life had the potential for variation, as opposed to the prevailing idea of his time that species could not change at all. The occasional appearance of wild-type pigeons and striped horses served him well in illustrating his argument. However, the concept of reversions wrongly assumes that a population will reveal its entire range of variation at one time. Some characteristics may not appear in the current generation, but may have existed in previous generations, or may be expressed in generations to come. All this shows is that variation exists both in space and time. In other words, although populations may show variation from one place to another, and from one generation to the next, the total amount of variation remains essentially the same.
Still, evolutionists see this wealth of living history as a storehouse of future change (e.g., Gould, 1983, pp. 185-186). Yet how can reversions contribute to the evolution of species, if contemporary forms keep regressing to a previous state? In his early years of research, Darwin admitted the possibility that “any great change in species is reduced by atavism” (see Barrett, et al., 1987, p. 259). So through time, a population may experience cycles of evolution and devolution without ever changing into a different species. Reversions may show how populations can vary, but they cannot prove large-scale change over long periods of time.
While older ideas on atavism persist in some textbooks, new research on embryos has changed the face of the argument (Gould, 1983). In one experiment, scientists used embryo tissue from a chick to stimulate the growth of teeth in embryonic mouse tissue. Evolutionists call this apparently hidden ability atavistic, supposing it to be a remnant of the chick’s tooth-bearing, reptilian ancestry. Also, the ability of chick and mouse tissues to work together is supposed to show their common ancestry, again, from a reptilian ancestor. Atavism, in this updated form, is not an additional argument for evolution, but a part of the argument from homology (i.e., that similar form implies similar ancestry).
The main problem with this interpretation is that embryonic cells are very flexible in their development. If chick and mouse tissues were taken from different places in the embryo, and then brought into contact, other body parts might develop. This experiment shows that embryonic cells can be manipulated to produce bizarre effects. Further, the similarity of embryonic cells that allowed these strange teeth to develop does not prove common ancestry. Rather, it is evidence of common design by an intelligent Creator Who used similar parts to achieve similar ends. Also, some fossil birds (like Archaeopteryx) had teeth. Perhaps God gave some bird kinds this ability, which they eventually lost through harmful mutations (Frair and Davis, 1983, p. 46).
EMBRYONIC RECAPITULATION
In the nineteenth century, Ernst Haeckel developed the theory of “embryonic recapitulation.” He believed that the biological development of an individual (ontogeny) repeats the evolutionary development of its species (phylogeny).
One oft-repeated “fact” of Haeckel’s theory is that mammalian embryos, including those of humans, develop “gill slits.” These are supposed to prove our evolutionary descent from fish. However, these “slits” are merely folds of flesh that later develop into the various parts of the neck region; they never function like the breathing organs of fish. For instance, although biologist Aaron Wasserman believes there are similarities between fish and mammal embryos at certain stages, he states that the mammalian embryo “can in no sense be called a fish; it never actually develops functional gills and is at all times a mammal” (1973, p. 497).
George Gaylord Simpson and William Beck also appear evasive on the issue of fish-like stages. In the body of their text they write:
Human embryos develop tails, which later disappear. They also develop gill-like pouches in the neck region, which disappear as such and are in part transformed into quite nongill-like structures, including the ear canal. At this stage the embryo looks a little like a fish, although the resemblance is not so close as is sometimes suggested (1965, p. 240).
Yet in a footnote they comment: “The human embryo does not have any differentiated gill tissue, and the gill-like pouches do not have open gill slits as in fishes. Fins are lacking. The tail is not at all like any fish’s tail. Indeed, the resemblance to an adult fish is vague and superficial” (1965, p. 240). The authors later conclude: “It is now firmly established that ontogeny does not recapitulate phylogeny” (1965, p. 241, emp. in orig.).
Further, Haeckel fabricated his drawings of embryos. W.R. Thompson makes the following comment:
When the “convergence” of embryos was not entirely satisfactory, Haeckel altered the illustrations of them to fit his theory. The alterations were slight but significant. The “biogenetic law” as a proof of evolution is valueless (1956, p. xvi).
Jane Oppenheimer adds other examples of fraud:
It was a failing of Haeckel as a would-be scientist that his hand as an artist altered what he saw with what should have been the eye of a more accurate beholder. He was more than once, often justifiably, accused of scientific falsification, by Wilhelm His and by many others. For only two examples, in Anthropogenie he drew the developing brain of a fish as curved, because that of reptiles, birds, and mammals is bent. But the vesicles of a fish brain always form a straight line. He drew the embryonic membranes of man as including a small sac-like allantois, an embryonic organ characteristic of and larger in reptiles, birds, and some nonhuman mammals. The human embryo has no sac-like allantois at all. Only its narrow stalk remains to conduct the umbilical blood vessels between embryo and placenta. Examples could be multiplied significantly (1988, p. 134).
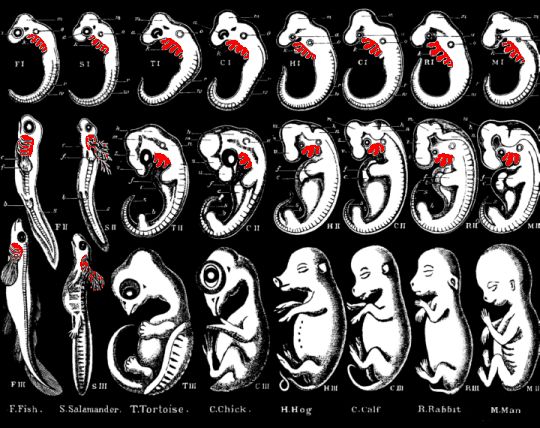 |
Haeckel’s drawings of embryos at three different stages for (from left to right): fish, salamander, tortoise, chick, hog, calf, rabbit and man (from 1876, Plates VI-VII). The supposed “gill-slits” are shown in red. |
Although Haeckel’s theory has fallen into disfavor, embryonic recapitulation has survived in a modified form. In this version, evolutionists argue that similar patterns of embryo development in different animals prove their common descent. For example, the neck folds in human embryos (which Haeckel labeled as “gill slits”) eventually give rise to the jaw. These folds develop from a similar area of the vertebrate column, and are controlled by similar gene sequences, as the gill arches of fish embryos (e.g., Gould, 1990, p. 16). The evolutionist concludes from this that fish and humans had a common ancestor.
However, this similarity is also evidence of common design. Mice, men, and pigs have four appendages and a head, and therefore it is not surprising that embryos should follow a similar path of development. Indeed, the ability of embryos to form in such perfection demands something more than evolution. Natural selection works primarily on organisms exposed to the environment or competition. Yet the embryo is isolated from the outside world. Evolutionary processes are at a loss to explain the origin of the mechanism that causes a fertilized egg to develop into a young version of the adult.
CONCLUSION
Although natural selection working on mutations remains the main thrust of evolutionary theory, the theory itself changes with the times. Many old ideas missed the mark, were unfounded, and in the worst cases, were deliberate falsehoods. Unfortunately, some continue to appear in science textbooks. It is still necessary to review the problems with these ideas, and how they have changed to accommodate new research.
The idea that animals carry diminished or useless parts is still popular among evolutionists. However, most of the rudimentary and vestigial structures they cite have real functions. Genuine cases of vestigial organs are rare, and may be the result of mutations. However, such changes are not sufficient to explain long-term, large-scale evolution.
Atavism and embryonic recapitulation were once put forward as separate lines of evidence in favor of evolution. Scientists now know that these arguments were faulty. Examples of atavism usually represent either natural variation or misunderstood birth defects. And contrary to the fraudulent work of Haeckel, human embryos never go through a fish, or any other nonhuman, stage. New twists on these ideas have arisen, but they reduce to the argument from homology. However, this is neither the only, nor the best, interpretation, because similarity also suggests design by an intelligent Creator.
REFERENCES
Barrett, Paul H., et al. (1987), Charles Darwin’s Notebooks, 1836-1844 (Ithaca, NY: Cornell University Press).
Bergman, Jerry and George Howe (1990), “Vestigial Organs” are Fully Functional (Terra Haute, IN: Creation Research Society).
Darwin, Charles (1859), The Origin of Species (New York: Avenel Books, 1979 reprint of the Penguin 1968 edition).
Frair, Wayne and Percival Davis (1983), A Case for Creation (Chicago, IL: Moody Press).
Gish, Duane T. (1983), “Evolution and the Human Tail,” Impact, No. 117.
Gould, Stephen Jay (1983), “Hen’s Teeth and Horse’s Toes,” Hen’s Teeth and Horse’s Toes (New York: W.W. Norton).
Gould, Stephen Jay (1990), “An Earful of Jaw,” Natural History, 99[3]:12-23, March.
Haeckel, Ernst (1876), The Evolution of Man (Akron, OH: Werner, translated from the German third edition).
Holland, Henry (1839), Medical Notes and Reflections (London). As quoted by Paul H. Barrett, et al. (1987).
Oppenheimer, Jane M. (1988), “Haeckel’s Variations on Darwin,” Biological Metaphor and Cladistic Classification, ed. H.M. Hoenigswald and L.F. Wiener (Pittsburgh, PA: University of Pennsylvania Press). As quoted in “Ernst Haeckel: Art vs. Science,” The SOR Bulletin, 1989, 5[1]:4.
Simpson, George G. and William S. Beck (1965), Life: An Introduction to Biology (New York: Harcourt, Brace & World).
W.R. Thompson (1956), “Introduction,” Origin of Species, by Charles Darwin (London: Dent, Everyman’s Library edition).
Wasserman, Aaron O. (1973), Biology (New York: Appleton-Century-Crofts).

REPRODUCTION & DISCLAIMERS: We are happy to grant permission for this article to be reproduced in part or in its entirety, as long as our stipulations are observed
posted by Believer | 3:00 AM
![]()
0 Comments:
Post a Comment
<< Home